


INTRODUCTION
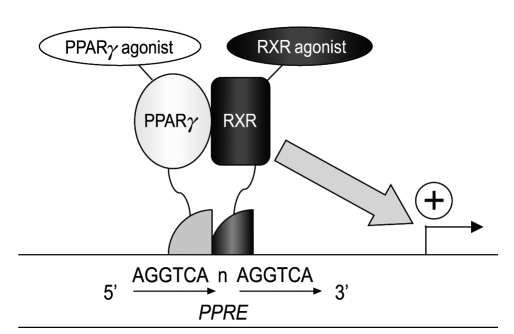
Peroxisome proliferator-activated receptor (PPAR)γ is a nuclear hormone receptor that, with the retinoid X receptor (RXR), binds as a heterodimer to the PPAR response element (PPRE), a direct repeat of 'AGGTCA' gapped by a nucleotide. PPAR is trans-activated by several agonists, including 15-deoxy-Δ12,14-prostaglandin J2 (15dPGJ2) and thiazolidinediones (TZDs); the latter are widely used as insulin-sensitizers in the treatment of diabetes [1] (Fig. 1). Recently, pleiotropic effects of PPARγ agonists in the vasculature have been demonstrated. These effects are independent of blood glucose-lowering activity and include protection against the progression of hypertension, atherosclerosis, and renal dysfunction [2]. In this review, we discuss recent findings regarding the additional beneficial aspects of PPARγ agonists in the vasculature, including conclusions based on our own data.
Effects of PPARg agonists in hypertension
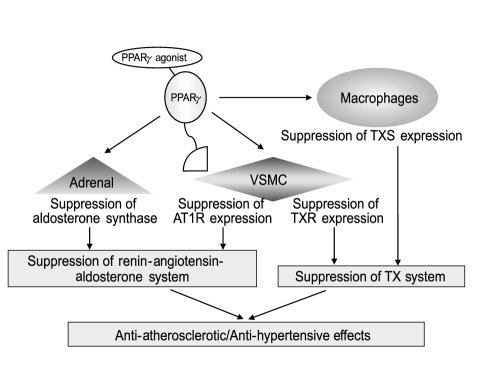
The blood-pressure-lowering effect of TZDs was recently demonstrated in a clinical study [4] and in the PROactive (PROspective pioglitAzone Clinical Trial In macroVascular Events) study, in which 5,238 type 2 diabetic patients were enrolled. Among the results of that trial, treatment with the TZD pioglitazone was shown to significantly decrease (3 mmHg) systolic blood pressure [5]. Because the renin-angiotensin (Ang)-aldosterone system (RAAS) plays the most important role in the progression of hypertension, we examined the effects of several PPARγ agonists on Ang II type 1 receptor (AT1R) expression in vascular smooth muscle cells (VSMCs). Interestingly, 15dPGJ2, as well as TZDs (pioglitazone, troglitazone, rosiglitazone), dose-dependently decreased the expression of AT1R mRNA [6,7].
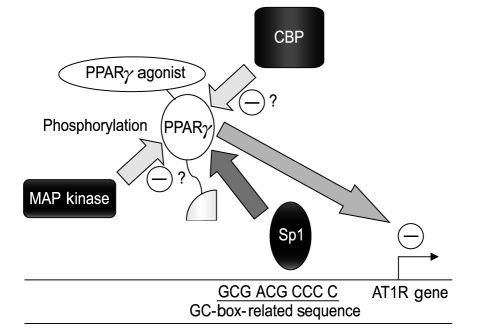
Transcriptional analysis using the rat AT1R gene promoter (-1969/+104) and AT1R mRNA stability analysis using actinomycin D together revealed that PPARγ agonists decrease AT1R expression at the transcriptional level. Mutation analysis of the promoter demonstrated that transcriptional suppression was mediated within the -58/-34 region (TGC AGA GCA GCG ACG CCC CCT AGG C) of the AT1R gene promoter, which contains a GC-box-related sequence (underlined), but lacks a PPRE [6] (Fig. 2). Instead, the transcription factor Sp1 was shown to bind to and trans-activate the promoter region [6]. Overexpression of PPARγ and Sp1, followed by transcriptional analysis, electrophoretic mobility shift assay, and glutathione S-transferase pull-down assay, revealed that agonist-activated PPARγ does not bind to the -58/-34 region, but rather to Sp1 via a protein-protein interaction [6]. Moreover, Sp1 binding to the region was inhibited by coincubation with PPARγ [6]. These results suggested that PPARγ-agonist-induced transcriptional suppression of the AT1R gene is mediated by the inhibition of Sp1 binding to the -58/-34 region through a protein-protein interaction between agonist-activated PPARγ and Sp1 (Fig. 2). Furthermore, transcriptional suppression was abrogated by the over-expression of co-activator CERB-binding protein (CBP) and PPARγ phosphorylation by mitogen-activated protein (MAP) kinase [8], most likely due to the functional modification of PPARγ (Fig. 2). PPARγ-agonist-mediated suppression of AT1R expression was also demonstrated in Ang-II-infused rats [9,10]. Moreover, PPARγ agonists have been shown to suppress Ang-II-induced phosphatidylinositol 3-kinase and MAP kinase [10] and to ameliorate Ang-II-mediated inflammatory responses by interfering with the Toll-like-receptor-4-dependent signaling pathway [11]. Additionally, using human adrenal H295R cells, we recently found an inhibitory effect of PPARγ agonists on Ang-II-induced aldosterone synthase expression and aldosterone secretion [12]. Thus, PPARγ agonists not only down-regulate AT1R expression but also inhibit Ang-II-mediated signaling pathways and adrenal aldosterone synthesis/secretion, which, together, may result in RAAS suppression (Fig. 3). The ability of PPARγ agonists to lower blood pressure has been reported in Ang-II-infused Sprague-Dawley rats [9,10], spontaneously hypertensive rats [13], deoxycorticosterone acetate-salt rats [14], and hypertensive double-transgenic mice expressing human renin and human angiotensinogen transgenes [15]. Conversely, transgenic mice expressing a dominant-negative PPARγ P465L mutation exhibited hypertension [16], consistent with the phenotype of patients with an equivalent PPARγ P467L mutation [17], without affecting RAAS components. Moreover, genetic manipulation of mice with varying PPARγ expression demonstrated that blood pressure was lowered by an increase in receptor expression and increased when levels of the receptor were reduced [18]. Taken together, these results suggest that the decrease in blood pressure mediated by PPARγ agonists occurs through several different mechanisms in addition to RAAS inhibition.
Effects of PPARγ agonists in protection against atherosclerosis
Thromboxane (TX) A2, which is generated from prostaglandin H2, stimulates the contraction and proliferation of VSMCs and may be involved in the progression of atherosclerosis. We thus examined the effect of PPARγ agonists on the expression of TX synthase (TXS) in macrophages [19] and the TX receptor (TXR) in VSMCs [7,20]. PPARγ agonist suppressed both TXS and TXR expression at the transcriptional level [7,19,20]. Detailed analysis revealed that agonist-activated PPARγ inhibited nuclear factor E2-related factor 2 (NRF2) binding to DNA of the TXS gene [19], and Sp1 binding to DNA of the TXR gene [20], in both cases via protein-protein interactions. Accordingly, PPARγ agonists may suppress the progression of atherosclerosis through inhibition of both the TX system, including the synthesis and action/signal-transduction function of TXA2 (Fig. 3), and RAAS.
Atherosclerosis is usually preceded by endothelial dysfunction, whereas PPARγ agonists have been reported to improve the function of these cells not only in streptozotocin-induced diabetic rats [21] and diabetic db/db mice [22], but also in type 2 diabetic patients [23] and non-diabetic patients with coronary artery disease [24]. Additionally, transgenic mice specifically expressing dominant-negative PPARγ in endothelium developed endothelial dysfunction in response to a high-fat diet [25]. PPARγ agonists have also been reported to reduce carotid intimal-medial thickness (CIMT) and in-stent restenosis after coronary intervention in diabetic and non-diabetic patients [26], neointima formation after balloon injury in rats [27], and instent restenosis in atherosclerotic rabbits [28]. Metaanalysis of controlled trials involving type 2 diabetic patients found a significant reduction in CIMT and pulse wave velocity by PPARγ agonists of the TZD group [29]. We examined the direct effect of PPARγ agonists on endothelial gene expression by performing DNA microarray analyses. In those experiments, confluent human umbilical vein endothelial cells (HUVEC) were treated for 24 hours with the TZD pioglitazone, at a concentration (100 nM) mimicking the serum concentration in patients after a single oral administration. RNA extracted from the cells was processed for DNA microarray analyses using Human Genome Oligo Set (Operon Biotechnologies Inc., Huntsville, AL, USA), allowing the analysis of approximately 35,000 genes. Representative regulated genes are shown in Table 1. Among the genes induced by pioglitazone were tissue inhibitor of metalloproteinases-3, prostacyclin receptor, kallikrein 6 and 11, prostaglandin E2 receptor (EP1 subtype), and microsomal glutathione Stransferase 3. Suppressed genes included matrix metalloproteinase-10 and plasminogen activator inhibitor-2 [30]. The protection of endothelial function by PPARγ agonists may thus proceed through the regulation of gene expression. Recently, PPARγ agonists were reported to stimulate endothelial nitric oxide (NO) production in HUVECs [31] and to increase the number and function of endothelial progenitor cells in patients with coronary artery disease [32]. Additionally, disruption of the endothelium-specific PPARγ in mice resulted in the reduction of vascular NO production without affecting endothelial NO synthase expression [33]. These observations, in addition to our DNA microarray findings, may also explain the antiatherogenic effects of PPARγ agonists.
Effects of PPARβ agonists in renal dysfunction
To examine the intra-renal localization of PPARγ protein, we generated an isoform-specific anti-PPARγ antibody, which was then used in the immunohistochemical analysis of Sprague-Dawley rat kidneys [34,35]. PPARγ protein was observed to be widely expressed in the nuclei of mesangial and epithelial cells in the glomeruli, proximal and distal tubules, loop of Henle, and medullary collecting ducts [34]. Additionally, the protein was detected in the intima/media of the renal vasculature [34]. We previously reported the vasodilating effects of the TZD troglitazone on the glomerular efferent arterioles of microdissected rabbit kidneys [36]. As suggested by the immunohistochemical data, these vasodilating effects may be mediated by PPARγ expressed in the intra-renal arterioles. The expression of PPARγ protein was also induced in distal tubules and cortical collecting ducts following administration of the TZD rosiglitazone to Sprague-Dawley rats [35]. These findings are potentially relevant in terms of pathophysiology, because TZDs have been reported to expand body fluid volume by the PPARγ-mediated stimulation of renal salt absorption through epithelial Na+ channels [37].
Renal protective effects of PPARγ ligands on type 2 diabetic patients with nephropathy, especially with respect to a reduction in urinary albumin, have recently been reported [38]. A meta-analysis of 15 studies involving 2,860 diabetic patients demonstrated a significant decrease in urinary albumin excretion in response to TZD-type PPARγ agonists [39]. Additionally, similar effects were observed in animal experiments using various rodent models of type 2 diabetes [38]. The mechanisms by which PPARγ agonists reduce urinary albumin remain unclear. However, together with their vasodilating effect on glomerular efferent arterioles [36], a lowering of blood pressure and an improvement of endothelial dysfunction may be cumulatively involved. Additionally, a recent study described the renal protective effect of PPARγ agonists against non-diabetic renal disease [40], indicating their general usefulness in the treatment of chronic kidney disease. We have also demonstrated a renal protective effect of the TZD rosiglitazone against cyclosporine-induced renal injury in Sprague-Dawley rats [41]. Moreover, the renal protective effect of the TZD pioglitazone against aging-related renal injury has been reported [42].
CONCLUSION
More than a decade has passed since the pleiotropic effects of PPARγ agonists were first reported. However, novel effects of PPARγ agonists are still being described on almost a monthly basis. In addition to the effects of these agents discussed in this review, anti-cancer activities of PPARγ agonist were recently reported [43]. We have also reported inhibitory effects of TZD-type PPARγ agonists on cell growth and REG (regenerating gene) Iα expression in gastrointestinal cancer cell lines [44]. It thus seems likely that the usefulness and effectiveness of PPARγ agonists against lifestyle-related diseases will be increasingly appreciated. This, in turn, may lead to further approved clinical applications of PPARγ agonists in the treatment of hypertension, atherosclerosis, and renal dysfunction, in addition to diabetes.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print





