


Decreased frequency and inflammatory change of FoxP3+ regulatory T cells in immunopathogenesis of human acute graft-versus-host disease
-
Nayeon Seong1,2,*, Cheongin Yang2,*, Kunye Kwak1, Ka-Won Kang1, Yong Park1, Byung Soo Kim1, Seong Hyun Jeong2, Joon Seong Park2, Yoon Seok Choi1,2

- Received January 12, 2025; Revised March 9, 2025; Accepted March 18, 2025;
- Abstract
-
- Background/Aims
- Acute graft-versus-host disease (GvHD) is a severe complication of allogeneic stem cell transplantation, characterized by immune-mediated tissue damage primarily affecting the skin, liver, and gastrointestinal tract. Regulatory T (Treg) cells play a critical role in maintaining immune homeostasis. However, the pathogenic roles of changes in Treg cell number and function on acute GvHD remain poorly understood. This study aimed to investigate the quantitative and qualitative changes in Treg cells and their clinical and pathogenic implication of acute GvHD.
- Methods
- A total of 62 patients who underwent allogeneic stem cell transplantation at a tertiary institution from 2019 to 2024 were enrolled. Peripheral blood mononuclear cells were isolated and analyzed by multicolor flow cytometry. Treg cell subsets and cytokine production were assessed after T-cell receptor stimulation. Serum levels of inflammatory cytokines were measured using cytometric bead array, and Treg cell suppressive function was evaluated through co-culture experiments.
- Results
- Patients with acute GvHD showed a decreased frequency of circulating Treg cells, with a notable increase in the CD45RA-FoxP3lo pro-inflammatory subset. Treg cells produced inflammatory cytokines including TNF-α upon stimulation and exhibited reduced suppressive activity. The frequency of TNF-α+ Treg cells correlated with the clinical severity of acute GvHD. Elevated serum levels of IL-6 and IL-21 were associated with the inflammatory conversion of Treg cells.
- Conclusions
- During human acute GvHD, frequencies of circulating Treg cells are significantly decreased. Inflammatory change of Treg cells, represented by TNF-α production and reduced suppressive capacity, contributes to the immunopathogenesis of acute GvHD.
- Graphical abstract
- Graphical abstract
- INTRODUCTION
- INTRODUCTION
Acute graft-versus-host disease (GvHD) remains a significant and potentially life-threatening complication following allogeneic hematopoietic stem cell transplantation (SCT) [1]. Acute GvHD is triggered by allo-reactive donor-derived T cells recognizing the recipient’s tissues, initiating an immune response that results in widespread tissue damage. The primary target organs affected by acute GvHD include the skin, liver, and gastrointestinal tract, leading to a range of clinical manifestations such as skin rash, jaundice, and severe diarrhea. Despite advancements in transplant techniques and immunosuppressive therapeutic strategies, acute GvHD continues to be a major challenge to the successful allogeneic SCT, contributing to considerable morbidity and mortality in transplant recipients [2].The pathophysiology of acute GvHD involves a complex interplay between donor T cells, host antigen-presenting cells (APCs), and pro-inflammatory cytokines [3,4]. Upon transplantation, donor T cells become activated by recognizing host allo-antigens presented by APCs, resulting in a cascade of various inflammatory cytokine production [5]. This inflammatory response further amplifies the immune response, causing immune-mediated tissue damage.Regulatory T (Treg) cells, a specialized subset of CD4+ T cells characterized by the expression of CD25 and the transcription factor FoxP3, play a crucial role in controlling excessive and inappropriate immune responses [6,7]. Although numerous studies have reported the potential role of Treg cells in the development of acute GvHD, the precise mechanisms underlying the pathologic changes of Treg cell population in this context remain unclear. Treg cells typically suppress the activation and proliferation of conventional T cells, thereby preventing excessive immune responses and maintaining immune homeostasis [8,9]. However, under inflammatory conditions, Treg cells can lose their suppressive function and acquire pro-inflammatory properties, a phenomenon known as “inflammatory conversion.” The inflammatory conversion of Treg cells has been observed in various autoimmune and inflammatory diseases, including multiple sclerosis, rheumatoid arthritis and psoriasis [10-12]. Also, previous study from our group has shown that Treg cells exhibit lineage plasticity, acquiring Th17-like characteristics during acute viral infections, with these inflammatory changes correlating with disease severity [13]. However, the impact of inflammatory conversion of Treg cells on the pathogenesis of acute GvHD has not yet been fully elucidated.In the present study, we aimed to investigate the quantitative and qualitative changes in Treg cells in patients with acute GvHD, especially focusing on their inflammatory conversion. We also analyze the association between the inflammatory change of Treg cells and clinically documented tissue damage.
- METHODS
- METHODS
- Study subjects and insolation of lymphocytes
- Study subjects and insolation of lymphocytes
This study was conducted from 2019 to 2024 and included adult patients (19 years or older) who underwent allogeneic SCT at Ajou University Medical Center for hematologic malignancies. Among the 154 patients who underwent allogeneic hematopoietic SCT during the study period, 40 patients diagnosed with acute GvHD were included in this study. The 22 patients in the control group who did not develop acute GvHD were selected based on matched clinical characteristics with the acute GvHD group to ensure comparability between the two groups. All Patients received peripheral blood stem cell transplants from matched sibling, matched unrelated, or haploidentical donors. For matched donor transplantation, the GvHD prophylaxis regimen consisted of a calcineurin inhibitor plus methotrexate and anti-thymocyte globulin [14]. For haploidentical donor transplantation, a calcineurin inhibitor and mycophenolate mofetil were administered in combination with post-transplant cyclophosphamide [15]. The severity of acute GvHD was assessed according to the Mount Sinai Acute GvHD International Consortium (MAGIC) criteria [16]. Detailed characteristics of 62 enrolled patients are described in Table 1. The study was approved by the Institutional Review Board of Ajou University Hospital (IRB number. AJOUIRB-OBS-2021-484), and all participants provided written informed consent. Additionally, all experimental procedures involving patient-derived specimens were conducted in accordance with the approved IRB protocol.For patients diagnosed with acute GvHD, peripheral blood samples were collected at the time of diagnosis corresponding to the respective grades of acute GvHD. The median time of sampling was day 56 post-transplantation (range, 17 to 89 days). For patients in the control group who did not develop acute GvHD, blood samples collected on day 35 post-transplantation were used. Peripheral blood mononuclear cells (PBMCs) were isolated from whole blood by standard Ficoll-Paque (GE Healthcare, Uppsala, Sweden) density gradient centrifugation. Serum samples were also obtained for cytokine analysis.- Flow cytometry and immunophenotyping
- Flow cytometry and immunophenotyping
The following fluorochrome-conjugated monoclonal antibodies used for multicolor flow cytometry: anti-CD3 (UCHT- 1), anti-CD4 (RPA-T4), anti-CD14 (M5E2), anti-CD19 (HIB19), anti-CD25 (M-A251), anti-CD45RA (HI100) (all from BD Biosciences, San Jose, CA, USA); anti-CD39 (eBioA1) (eBioscience, San Diego, CA, USA); anti-CD127 (40131) (R&D Systems, Minneapolis, MN, USA). Cryopreserved or freshly isolated lymphocytes were stained with fluorochrome-conjugated antibodies against surface markers for 30 minutes on ice and then washed. In all samples for flow cytometric analyses, dead cells were excluded by using Live/Dead fixable cell stain kit (Invitrogen, Carlsbad, CA, USA).For intracellular staining, PE-conjugated anti-Foxp3 monoclonal antibody (PCH101) and anti-Helios (22F6) (all from eBioscience). Surface-stained cells were permeabilized using FoxP3 staining buffer kit (eBioscience) according to the manufacturer’s instruction and further stained for intracellular molecules.Flow cytometry was performed on an LSR II instrument using FACSDiva software (BD Biosciences), and the data were analyzed using FlowJo software (Treestar, San Carlos, CA, USA).- In vitro stimulation of T cells and intracellular cytokine staining
- In vitro stimulation of T cells and intracellular cytokine staining
Cryopreserved PBMCs were thawed, resuspended in RPMI 1640 containing 5% fetal bovine serum (FBS) and 2 mM L-glutamine, and rested overnight at 37°C. PBMCs were stimulated in the presence of anti-CD3 antibody (0.1 μg/mL) and anti-CD28 antibody (1 μg/mL), and brefeldin A (GolgiPlug; BD Biosciences) and monensin (GolgiStop; BD Biosciences) were added 1 hour later. After another 5 hours of incubation, PBMCs were first stained with Live/Dead fixable cell stain kit to exclude dead cells and then stained with fluorochrome-conjugated antibodies against surface markers. Stained cells were permeabilized using FoxP3 staining buffer kit and further stained for intracellular molecules with anti-FoxP3, either anti-IFN-γ (BD Biosciences) or anti-IL-17A (eBioscience) and anti-TNF-α (BD Biosciences).- Standard Treg cell suppression assay
- Standard Treg cell suppression assay
The suppressive function of Treg cells was assessed using co-culture experiments with CFSE-labeled responder T cells as described elsewhere. Briefly, CFSE-labeled Treg-depleted responder CD4+ T cells (Tresp) were cocultured with magnetically isolated Treg cells in a range of ratios as indicated within RPMI 1640 containing 5% FBS and 2 mM L-glutamine. Treg cells are isolated from PBMCs using CD4+CD25+CD127lo/- regulatory T cell isolation kit II (Miltenyi Biotec, Bergisch Gladbach, Germany) according to manufacturer’s protocol. At the end of coculture in the presence of soluble anti-CD3 anti-body (0.1 μg/mL; Beckman Coulter Immunotech, Marseille, France) and anti-CD28 antibody (1 μg/mL; BD Biosciences) stimulation for 96 hours, cells were analyzed for proliferation assessed by percentage of CFSElo dividing cells. Percentage of suppression was calculated as [1 - (% T cell proliferation with Treg cells / % T cell proliferation without Treg cells)] × 100.- Serum cytokine analysis
- Serum cytokine analysis
Levels of soluble factors including IL-6 and IL-21 in sera of acute GvHD patients were quantified using CBA (BD Biosciences). Briefly, capture beads and PE-conjugated detection reagent for each soluble factor analyzed were incubated with serum of patients. Concentration in sera was determined by measuring fluorescence intensity of beads capturing soluble factors with LSRII flow cytometer and interpolating the value into standard equation.- Statistical analyses
- Statistical analyses
Statistical analyses were performed using the Prism software version 5.0 (GraphPad, La Jolla, CA, USA). The Mann–Whitney U test was used for comparisons between two groups, and the Wilcoxon matched-pairs signed-rank test was used for paired values. A p value ≤ 0.05 was considered statistically significant.
- RESULTS
- RESULTS
- Decreased frequency of circulating Treg cells during acute GvHD
- Decreased frequency of circulating Treg cells during acute GvHD
We first determined the frequency of Tregs in the peripheral blood of patients with acute GvHD, which was collected at the time of diagnosis of GvHD. Flow cytometric analysis revealed a significant reduction in the percentage of CD25hiCD127lo/-FoxP3+ CD4+ Treg cells in patients with acute GvHD compared to non-acute GvHD control (Fig. 1A). We also investigated the association between quantitative and qualitative changes in Treg cells and clinical factors that could influence acute GvHD. However, no significant associations were identified. The human CD4+CD25hiFoxp3+ T cell population is known to be functionally heterogenous and composed of three distinct subpopulations such as CD45RA+Foxp3lo resting Tregs, CD45RA-Foxp3hi activated Tregs and cytokine-secreting CD45RA-Foxp3lo non-suppressive cells. When we analyzed for the composition of Treg cell subsets in more detail, we observed that the proportion of CD45RA-FoxP3hi (actively suppressive) Treg cells was decreased and the proportion of CD45RA-FoxP3lo (cytokine-producing non-suppressive) Treg cells was increased in patients with acute GvHD (Fig. 1B). Proportion of CD45R+FxoP3+ (naive) Treg cells was also decreased in patients with acute GvHD (Fig. 1B).- CD4+CD25hiCD127lo/-FoxP3+ Treg cells from acute GvHD patients produce inflammatory cytokine TNF-α
- CD4+CD25hiCD127lo/-FoxP3+ Treg cells from acute GvHD patients produce inflammatory cytokine TNF-α
We assessed the production of inflammatory cytokines from Treg cells following TCR stimulation of peripheral blood lymphocytes with anti-CD3/CD28 antibody and intracellular cytokine staining. As a result, we found that a remarkable proportion of CD4+CD25hiFoxP3+ Treg cells of acute GvHD patients produced TNF-α upon TCR stimulation (Fig. 2A). Notably, a significant proportion of TNF-α-producing Treg cells also secreted IL-17A simultaneously (Fig. 2A). These findings align with the increased CD45RA-FoxP3lo fraction observed among Treg cell subsets in aGvHD patients (Fig. 1B).To further explore the clinical implications of this inflammatory Treg cell population, we analyzed their correlation with the progression and severity of acute GvHD. The frequency of TNF-α+ cells among circulating Treg cells significantly correlated with the severity of acute GvHD, with higher proportion of TNF-α+ Treg cells in patients with the higher grade of acute GvHD (Fig. 2B). This suggests that the pathological conversion of Treg cells to a pro-inflammatory state, characterized by TNF-α production, contributes to immune-mediated tissue injury during acute GvHD.- Treg cell suppressive function is reduced during acute GvHD
- Treg cell suppressive function is reduced during acute GvHD
Next, we evaluated immunophenotypes known to be related to Treg cell suppressive function using multicolor flow cytometry. Our analysis revealed that TNF-α+ Treg cells of acute GvHD patients exhibited lower expression of FoxP3, a master transcriptional factor of Treg cell function, compared to the TNF-α- counterpart (Fig. 3A). Similarly, the proportion of CD39+ cells, a marker associated with Treg cell suppressive activity, was reduced in TNF-α+ Treg cells relative to TNF-α- cells (Fig. 3B). Additionally, we observed a decreased frequency of Helios-expressing Treg cells, a key determinant of Treg cell suppressive capacity, in TNF-α+ Treg cells from acute GvHD patients (Fig. 3B).We further performed Treg cell suppression assay to quantify their functional capacity on a per-cell basis, using peripheral blood T cells of recipient of allogeneic SCT both at baseline and post-acute GvHD onset. Notably, the suppressive activity of Treg cells during acute GvHD was significantly diminished compared to pre-SCT controls, as evidenced by reduced inhibition of non-Treg CD4+ T cell proliferation in co-culture experiments (Fig. 3C). These findings indicate that the impaired suppressive function of Treg cells, coupled with their inflammatory conversion, may contribute to the immunopathogenesis of acute GvHD.- Th17-polarizing cytokines are abundant in the serum of acute GvHD patients
- Th17-polarizing cytokines are abundant in the serum of acute GvHD patients
To investigate a potential mechanism underlying inflammatory changes in Treg cells, we quantified various soluble factors in systemic circulation before and after onset of acute GvHD. Our analysis demonstrated that serum levels of IL-6 (Fig. 4A) and IL-21 (Fig. 4B) were significantly elevated following the onset of acute GvHD compared to baseline levels. These findings suggest that these Th17-polarizing cytokines may play a pivotal role in driving the inflammatory conversion of Treg cells during the course of acute GvHD, thereby contributing to immune dysregulation and tissue injury.
- DISCUSSION
- DISCUSSION
Our study underscores the critical role of the inflammatory conversion of Treg cells in the pathogenesis of human acute GvHD. Beyond a reduction in the frequency of circulating Treg cells, these cells undergo functional changes, acquiring the ability to produce pro-inflammatory cytokines such as TNF-α and IL-17A, which significantly contribute to immune-mediated tissue damage. Furthermore, these inflammatory Treg cells demonstrated diminished suppressive activity and were strongly correlated with the clinical severity of acute GvHD, highlighting their pathogenic roles.Many studies have demonstrated that the frequency of peripheral blood Treg cells is inversely associated with tissue inflammation in various autoimmune disorders [17]. Similarly, our findings showed a reduction in the number of circulating Treg cells at the onset of acute GvHD. However, given that human Treg cells comprise heterogeneous functional subpopulations, the absolute number of total Treg cells may not fully reflect their pathophysiological significance [18-20]. In this study, we conducted a detailed analysis of three distinct functional subsets of Treg cells. Specifically, we observed a decrease in the proportion of CD45RA-FoxP3hi Treg cells, known for their strong suppressive activity, alongside an increase in CD45RA-FoxP3lo Treg cells, which exhibit cytokine-producing, pro-inflammatory characteristics. These shifts in Treg cell subpopulations suggest that the balance between suppressive and pro-inflammatory Treg cells plays a pivotal role in the progression of acute GvHD.The mechanisms underlying the functional alterations of Treg cells during acute GvHD remain poorly understood. It has been generally hypothesized that the inflammatory environment present in various immune disorders contributes to these unexpected functional changes [21,22]. In this study, we aimed to identify potential drivers of the inflammatory conversion of Treg cells and propose that elevated levels of pro-inflammatory cytokines, such as IL-6 and IL-21, found in the peripheral blood of acute GvHD patients, may play a crucial role. Both IL-6 and IL-21 are well-known for promoting the differentiation of naïve CD4+ T cells into the Th17 lineage in both in vitro and in vivo settings, resulting in IL-17A production and subsequent tissue inflammation [23]. Consistent with this, our study documented that TNF-α-producing Treg cells also produced IL-17A, indicating a simultaneous acquisition of multiple pro-inflammatory properties during acute GvHD.In addition to uncovering the underlying mechanisms of Treg cell conversion, our findings suggest that elevated levels of Th17-polarizing cytokines may have significant implications for the development of novel therapeutic strategies against acute GvHD. Targeting these cytokines and preventing the inflammatory conversion of Treg cells could help mitigate the severity of acute GvHD. In clinical practice, monitoring the frequency and phenotype of inflammatory Treg cells could serve as a valuable biomarker for predicting acute GvHD severity and treatment response [24]. Furthermore, therapeutic approaches aimed at stabilizing Treg cell function or preventing their conversion into pro-inflammatory cells have the potential to significantly improve clinical outcomes for patients with acute GvHD [25,26].Inflammatory conversion represents a broader paradigm of immune regulation, with relevance to a variety of immune- mediated disorders. Future research should explore additional molecular mechanisms affecting Treg cell plasticity and stability. A deeper understanding of Treg cell plasticity and inflammatory conversion could pave the way for targeted interventions that enhance immune regulation while preventing pathological immune responses across multiple immune-mediated conditions.
- KEY MESSAGE
- KEY MESSAGE
1. During human acute GvHD, frequencies of circulating Treg cells are significantly decreased.2. Treg cells undergo inflammatory conversion, producing pro-inflammatory cytokines TNF-α and exhibiting reduced suppressive activity.3. This conversion is closely associated with clinical severity, suggesting a role in the immunopathogenesis of acute GvHD.
- Notes
- Notes
-
CRedit authorship contributions Nayeon Seong: investigation, data curation, writing - original draft; Cheongin Yang: data curation, formal analysis, writing - original draft; Kunye Kwak: data curation, formal analysis; Ka-Won Kang: data curation, formal analysis; Yong Park: data curation, formal analysis, writing - review & editing; Byung Soo Kim: data curation, formal analysis; Seong Hyun Jeong: data curation, formal analysis; Joon Seong Park: writing - review & editing, supervision; Yoon Seok Choi: conceptualization, methodology, writing - original draft, writing - review & editing, supervision, funding acquisition
- Conflicts of Interest
- Conflicts of Interest
-
Conflicts of interest The authors disclose no conflicts.
- Notes
- Notes
-
Funding This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korea government (Ministry of Science and ICT) (NRF-2018R1C1B6008261 and RS-2022-NR070625 to Y.S.C.).
Figure 1.
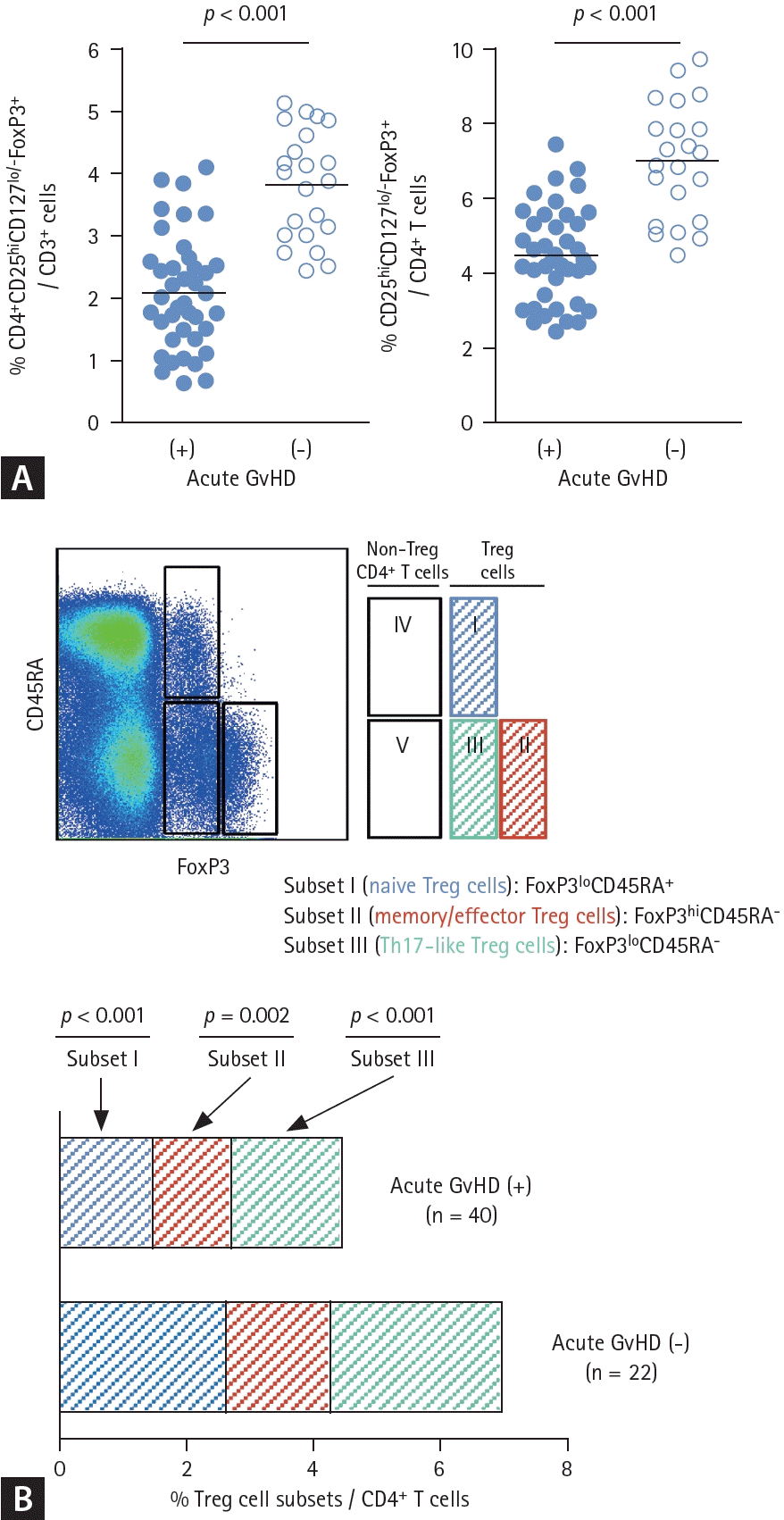
Figure 2.
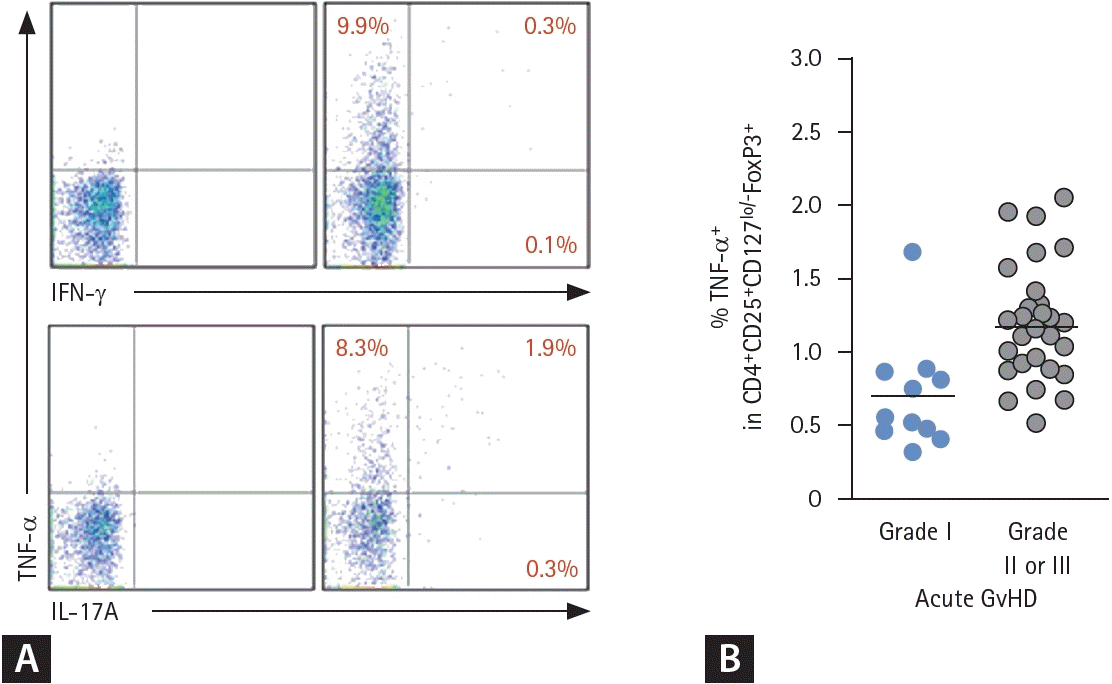
Figure 3.
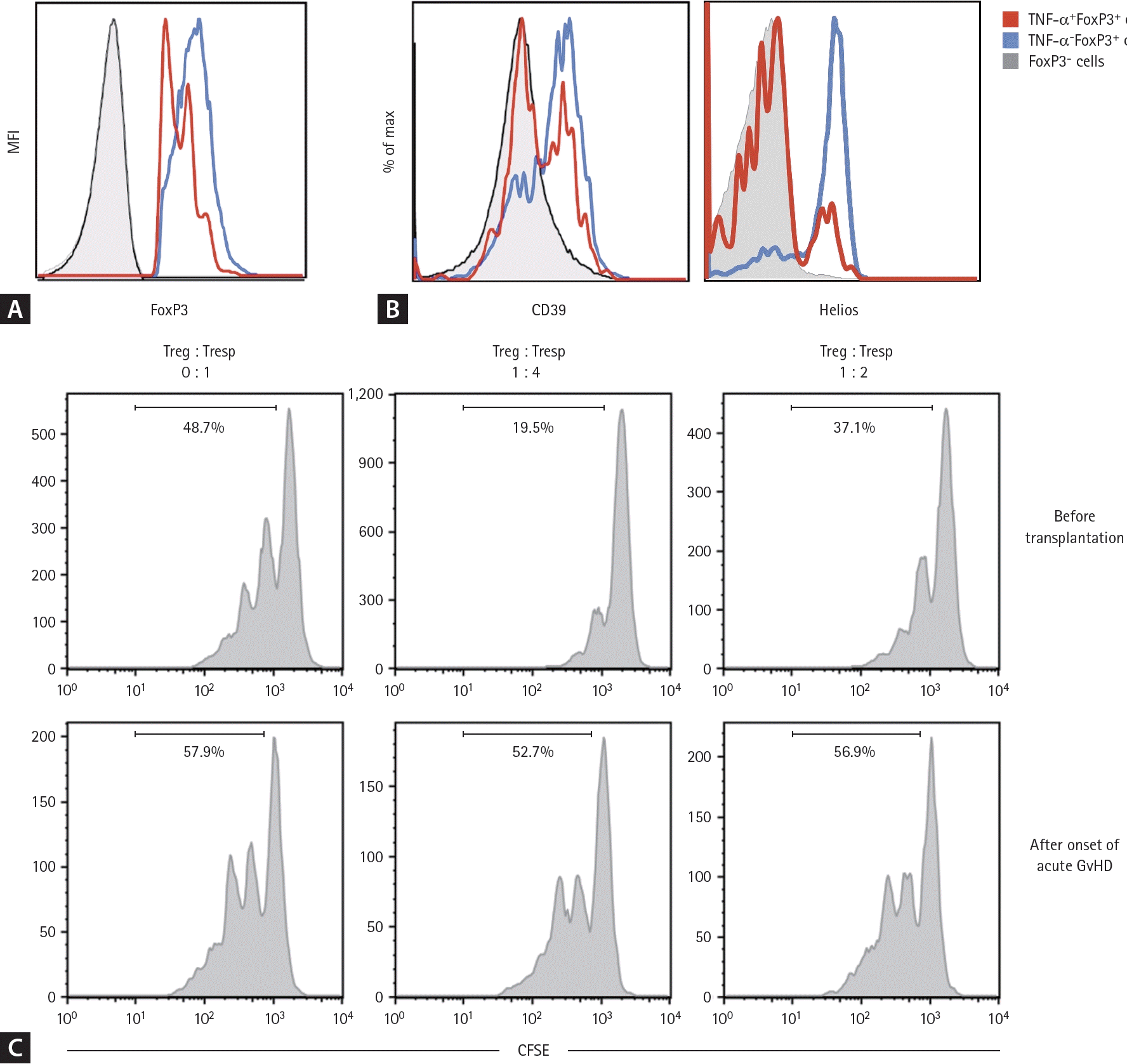
Figure 4.
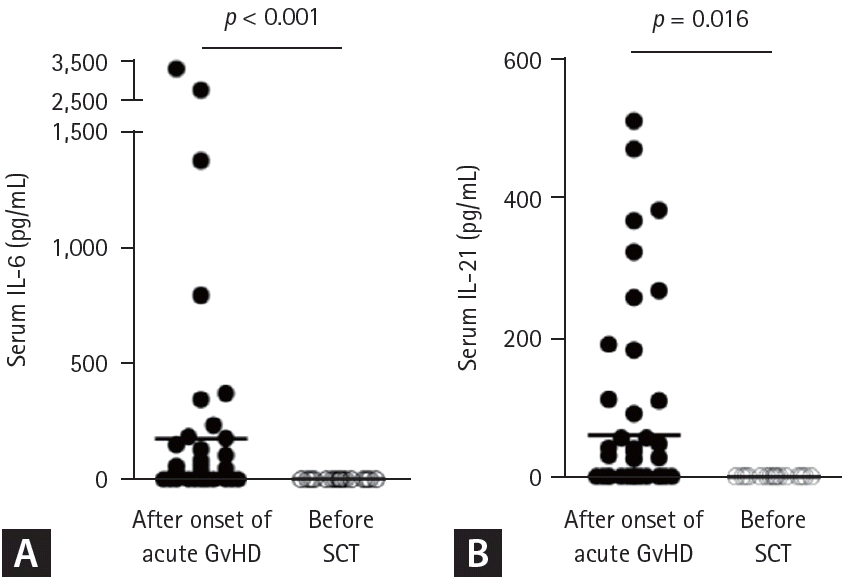
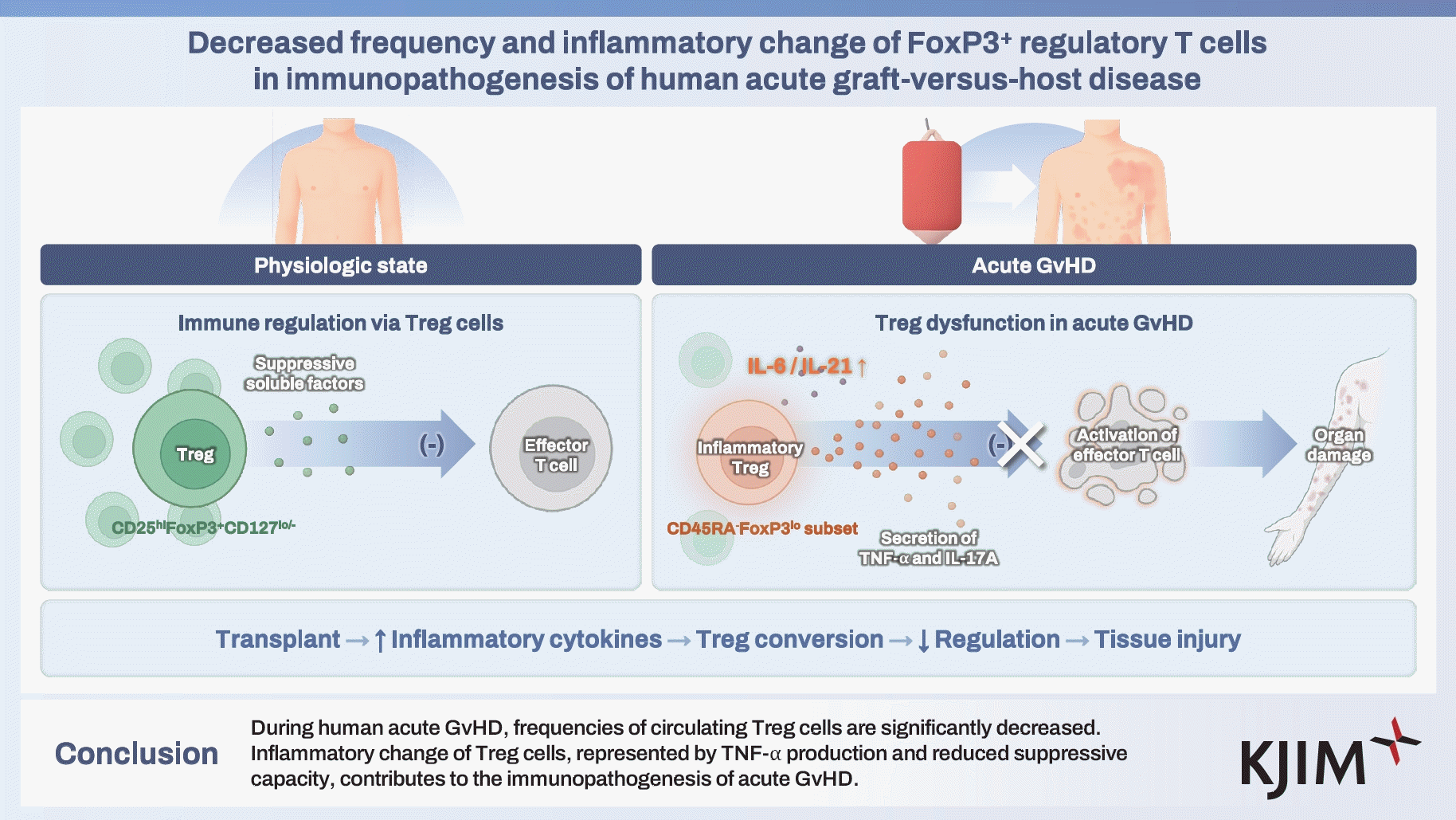
Table 1.
- References
- References
REFERENCES
1. Granot N, Storb R. History of hematopoietic cell transplantation: challenges and progress. Haematologica 2020;105:2716–2729.
[Article] [PubMed] [PMC]2. Kean LS, Blazar BR. Major breakthroughs in hematopoietic stem cell transplantation and future challenges in clinical implementation. J Clin Invest 2024;134:e179944.
[Article] [PubMed] [PMC]3. Zeiser R, Blazar BR. Acute graft-versus-host disease - biologic process, prevention, and therapy. N Engl J Med 2017;377:2167–2179.
[Article] [PubMed] [PMC]4. Hill GR, Betts BC, Tkachev V, Kean LS, Blazar BR. Current concepts and advances in graft-versus-host disease immunology. Annu Rev Immunol 2021;39:19–49.
[Article] [PubMed] [PMC]5. Zeiser R. Advances in understanding the pathogenesis of graft-versus-host disease. Br J Haematol 2019;187:563–572.
[Article] [PubMed]6. Ohkura N, Kitagawa Y, Sakaguchi S. Development and maintenance of regulatory T cells. Immunity 2013;38:414–423.
[Article] [PubMed]7. Dikiy S, Rudensky AY. Principles of regulatory T cell function. Immunity 2023;56:240–255.
[Article] [PubMed]8. Sakaguchi S. Taking regulatory T cells into medicine. J Exp Med 2021;218:e20210831.
[Article] [PubMed] [PMC]9. Morina L, Jones ME, Oguz C, et al. Co-expression of Foxp3 and Helios facilitates the identification of human T regulatory cells in health and disease. Front Immunol 2023;14:1114780.
[Article] [PubMed] [PMC]10. Bovenschen HJ, van de Kerkhof PC, van Erp PE, Woestenenk R, Joosten I, Koenen HJ. Foxp3+ regulatory T cells of psoriasis patients easily differentiate into IL-17A-producing cells and are found in lesional skin. J Invest Dermatol 2011;131:1853–1860.
[Article] [PubMed]11. Dominguez-Villar M, Baecher-Allan CM, Hafler DA. Identification of T helper type 1-like, Foxp3+ regulatory T cells in human autoimmune disease. Nat Med 2011;17:673–675.
[Article] [PubMed] [PMC]12. Wang T, Sun X, Zhao J, et al. Regulatory T cells in rheumatoid arthritis showed increased plasticity toward Th17 but retained suppressive function in peripheral blood. Ann Rheum Dis 2015;74:1293–1301.
[Article] [PubMed]13. Choi YS, Jung MK, Lee J, et al. Tumor necrosis factor-producing T-regulatory cells are associated with severe liver injury in patients with acute hepatitis A. Gastroenterology 2018;154:1047–1060.
[Article] [PubMed] [PMC]14. Lee JM, Kim DY, Cho HJ, et al. Reduced-intensity chemotherapy with tyrosine kinase inhibitor followed by allogeneic transplantation is effective in patients with Philadelphia chromosome-positive acute lymphoblastic leukemia. Korean J Intern Med 2025;40:124–134.
[Article] [PubMed] [PMC]15. Hong KT, Kim BK, An HY, et al. Comparing haploidentical transplantation with post-transplantation cyclophosphamide and umbilical cord blood transplantation using targeted busulfan in children and adolescents with hematologic malignancies. Blood Res 2025;60:7.
[Article] [PubMed] [PMC]16. Harris AC, Young R, Devine S, et al. International, multicenter standardization of acute graft-versus-host disease clinical data collection: a report from the Mount Sinai Acute GVHD International Consortium. Biol Blood Marrow Transplant 2016;22:4–10.
[Article] [PubMed] [PMC]17. Rodríguez-Gil A, Pérez-Simón JA, Ritz J, Lacerda JF, Soares MV. Editorial: regulatory T cells in graft versus host disease. Front Immunol 2023;13:1085220.
[PubMed] [PMC]18. Miyara M, Yoshioka Y, Kitoh A, et al. Functional delineation and differentiation dynamics of human CD4+ T cells expressing the FoxP3 transcription factor. Immunity 2009;30:899–911.
[Article] [PubMed]19. Choi YS, Lee J, Lee HW, et al. Liver injury in acute hepatitis A is associated with decreased frequency of regulatory T cells caused by Fas-mediated apoptosis. Gut 2015;64:1303–1313.
[Article] [PubMed]20. Wing JB, Tanaka A, Sakaguchi S. Human FOXP3+ regulatory T cell heterogeneity and function in autoimmunity and cancer. Immunity 2019;50:302–316.
[Article] [PubMed]21. Miyao T, Floess S, Setoguchi R, et al. Plasticity of Foxp3+ T cells reflects promiscuous Foxp3 expression in conventional T cells but not reprogramming of regulatory T cells. Immunity 2012;36:262–275.
[Article] [PubMed]22. Saini C, Sapra L, Bhardwaj A, et al. IL-21 plays an important role in modulating “Th17-Treg” cell axis in leprosy Type 1 reactions. Cytokine 2022;152:155821.
[Article] [PubMed]23. Kimura A, Kishimoto T. IL-6: regulator of Treg/Th17 balance. Eur J Immunol 2010;40:1830–1835.
[Article] [PubMed]24. Bidgoli A, DePriest BP, Saatloo MV, Jiang H, Fu D, Paczesny S. Current definitions and clinical implications of biomarkers in graft-versus-host disease. Transplant Cell Ther 2022;28:657–666.
[Article] [PubMed] [PMC]