


INTRODUCTION
Asthma, a chronic airway inflammatory disease, is an important source of morbidity and mortality worldwide [1]. Current guidelines recommend corticosteroid treatment for the management of asthma [2-4]. Endogenous corticosteroid levels and therapeutic responses to exogenous corticosteroids are known to be influenced by genetics, with heritability ranged from 0.40 to 0.56 [5-7]. This suggests a pharmacogenetic basis for the intraindividual variability of corticosteroid responsiveness in asthmatics [8].
Histone deacetylase (HDAC) plays a key role in the regulation of inflammatory genes by removing acetyl groups from histones [9,10]. It has been reported that conditional deletion of HDAC1 in T cells resulted in enhanced airway inflammation and increased Th2 cytokine production [11]. Moreover, HDAC is involved in the mechanism of action of corticosteroids. For example, recruitment of HDAC2 to activated inflammatory genes is an important mechanism of inflammatory gene repression by corticosteroids and HDAC2 activity is reduced in some diseases in which patients showed poor responses to corticosteroids treatment [12]. Together, HDAC1 and HDAC2 may be target molecules to predict therapeutic responses to corticosteroids. However, to date, any genetic effects of HDAC1 and HDAC2 on the response to corticosteroids in asthmatics have remained unknown.
In the present study, we evaluated associations between asthma severity, which is related to reduced responsiveness to corticosteroids [13], and single nucleotide polymorphisms (SNPs) in HDAC1 and HDAC2 in adult asthmatics. Then, we further assessed the effects of those SNPs on responsiveness to corticosteroids, as measured in terms of the change in lung function, in independent adult and childhood asthmatics.
METHODS
All subjects enrolled in this study provided written informed consent. The study protocol was approved by the Institutional Review Board of Seoul National University Hospital (H-1110-130-384).
Primary analysis
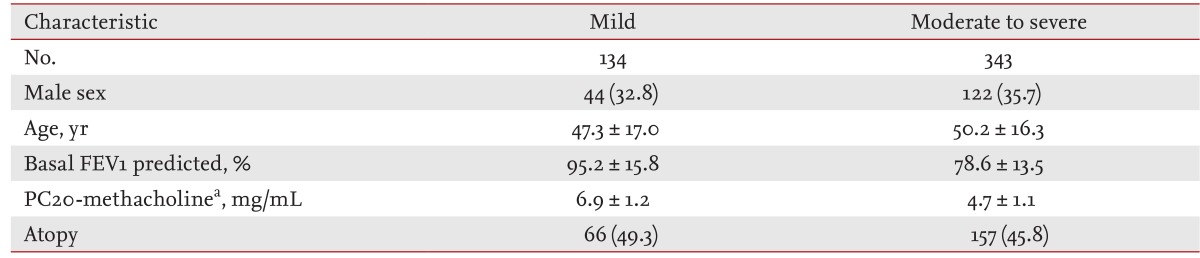
In total, 477 asthmatics were enrolled at Seoul National University Hospital, Seoul, Korea. The diagnosis of asthma was made at least 6 months prior to enrollment, according to current guidelines [2-4]: episodic symptoms including wheezing, coughing, and dyspnea plus a positive bronchodilator response (an increase in forced expiratory volume in 1 second [FEV1], from baseline that was more than 200 mL and more than 15% of the prebronchodilator value), or a provocative concentration of methacholine causing a 20% reduction in FEV1 (PC20) Ōēż 16 mg/mL [2-4]. Enrolled asthmatics were treated with conventional medications based on the The Global Initiative for Asthma guideline according to their asthma control status [2]. Asthma severity was determined based on lung function and the medication use index needed to obtain control, as described previously [14,15].
Secondary analysis
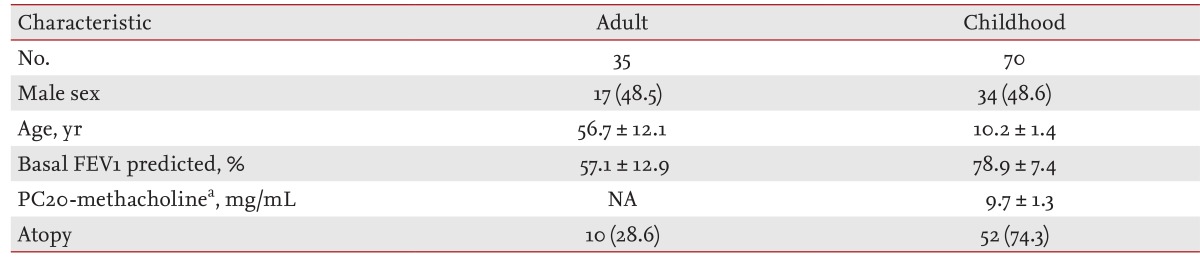
To generalize the results obtained in the primary analysis, we enrolled adult (treatment-na├»ve 35 subjects, systemic corticosteroid group) and childhood asthmatics (treatment-na├»ve 70 subjects, inhaled corticosteroid group) in a secondary analysis. Adult asthmatics visited our clinic first with dyspnea and decreased lung function (FEV1 Ōēż 80% predicted value) and were diagnosed with asthma based on positive bronchodilator responses. They were treated with short-term systemic steroids alone for prompt relief of their symptoms (oral prednisolone 15 mg, twice per day for 7 days). Childhood asthmatics enrolled at the Asan Medical Center, Seoul, Korea, were diagnosed with asthma according to current treatment guidelines [2-4]. All were treated with inhaled corticosteroids for 8 weeks according to their asthma control status. Treatment responses in both groups were measured by the degree of increase in FEV1: %FEV1 increase = (FEV1 after treatment - FEV1 at baseline)/FEV1 at baseline ├Ś 100.
Genotyping
The assessment of the SNPs in HDAC1 and HDAC2 was described in our previous study [16]. Briefly, after isolating genomic DNA from the peripheral blood of 24 healthy Korean subjects using the QIAamp DNA blood kit following the manufacturer's protocol (Qiagen, Hilden, Germany), we amplified 2 kb of the 5'-upstream region in the promoter and all exons, including the exon-intron boundaries, of HDAC1 and HDAC2 by polymerase chain reaction (PCR; reference genome sequences; NM_004964.2 [HDAC1] and NM_001527.3 [HDAC2]). Amplified PCR products were sequenced using a Big Dye Terminator Cycle Sequencing Ready Reaction Kit (Applied Biosystems, Foster City, CA, USA) in both directions according to standard protocols. After sequencing of HDAC1 and HDAC2, we identified five SNPs in HDAC1 and 14 SNPs in HDAC2 (Supplementary Tables 1 and 2). Among these, two SNPs (rs1741981 in HDAC1 and rs58677352 in HDAC2) were selected for scoring after considering minor allele frequencies (higher than 5%) and location (in exons and 5', near the gene). Scoring was conducted with the high throughput single base-pair extension method (SNP-IT assay) using an SNPstream25K system, which was customized to automatically genotype DNA samples in 384-well plates and to provide a colorimetric readout (Orchid Biosciences, Princeton, NJ, USA) as described previously [17].
Statistical analyses
An association analysis, based on a case-control design, was performed for each SNP using a genetic model approach: an allele model (A [major allele] vs. B [minor allele]), a dominant model (AA vs. AB + BB), and a recessive model (BB vs. AA + AB). Hardy-Weinberg equilibrium was tested using 2 ├Ś 2 tests. All statistical analyses were performed with the R version 2.15.3 (http://www.R-project.org/). p values of less than 0.05 were considered to indicate statistical significance.
RESULTS
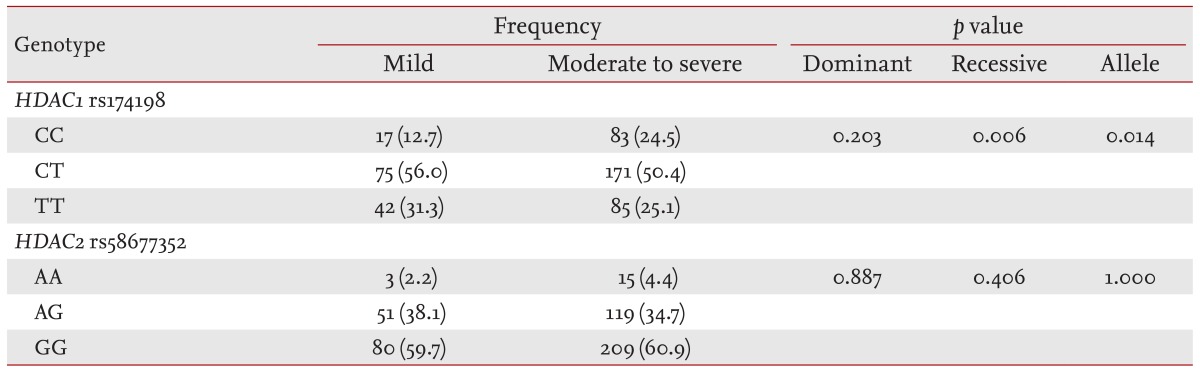
The characteristics of the asthmatics enrolled in the primary and secondary analyses are shown in Tables 1 and 2, respectively. Rs1741981 in HDAC1 and rs58677352 in HDAC2 were in Hardy-Weinberg equilibrium. The primary analysis revealed that rs1741981 was significantly related to asthma severity in a recessive model (p = 0.006) (Table 3). This relationship was still significant after the Bonferroni correction (multiplied by 6 [2 SNPs ├Ś 3 models]; corrected p = 0.036). However, rs58677352 showed no relationship with asthma severity (data not shown). As rs1741981 showed a significant relation with asthma severity in a recessive model, the secondary analysis was done using the same (recessive) model. Adult asthmatics with the CC genotype of rs1741981 (n = 12) showed significantly lower %FEV1 increases in response to systemic corticosteroid treatment compared with those with the CT or TT genotype (12.7% ┬▒ 7.2% vs. 37.4% ┬▒ 33.7%, p = 0.018) (Fig. 1A). The same tendency was observed in childhood asthmatics. Subjects with the CC genotype of rs1741981 (n = 15) showed significantly lower %FEV1 increases in response to inhaled corticosteroid treatment compared with those with the CT or TT genotype (14.1% ┬▒ 5.9% vs. 19.4% ┬▒ 8.9%, p = 0.035) (Fig. 1B). The combined p value, calculated from the one-sided p values of the secondary populations using Stouffer z-transform test [18], was 0.0019. However, rs58677352 showed no relationship with lung function improvement in response to corticosteroid treatment adults or childhood asthmatics (data not shown).
DISCUSSION
The present study demonstrated that rs1741981 in HDAC1 is significantly associated with the response to corticosteroid treatment in asthmatics. To our knowledge, this is the first report of a genetic association between HDAC1 and response to corticosteroids in asthmatics.
Corticosteroids are among the most effective treatments for asthma [19], and reduce inflammation via glucocorticoid receptor-mediated recruitment of HDAC2 [20]. Because reduced HDAC2 expression was observed in bronchial biopsies of asthmatic patients [21] and HDAC2-mediated deacetylation of the corticosteroid receptor enabled nuclear factor-╬║B suppression in airway epithelial cell line and alveolar macrophages [22], the discussion of the corticosteroid response in asthma has to date focused on HDAC2 alone. However, several investigators have argued that corticosteroid action in asthma is explained partially by inhibition of T cell activation [23-25]. Recently, it has been reported that that T cell-specific loss of HDAC1 resulted in enhanced allergic airway inflammation, by modulating cytokine production [11]. Moreover, HDAC1 was localized within most airway cells and infiltrating inflammatory cells in the lung in a murine asthma model, and the HDAC inhibitor TSA attenuated allergic airway inflammation in mice by reducing T cell infiltration and Th2 cytokine production [26]. Taken together, HDAC1 may play an important role in mediating the anti-inflammatory action of corticosteroids. Accordingly, our data provide pharmacogenetic evidence that a genetic polymorphism in HDAC1 can predict the response to corticosteroid treatment in asthmatics.
Functional variants strengthen the reliability of results of a pharmacogenetic study. Rs1741981 is located in the 5' near gene region of HDAC1. We used a web-based tool to evaluate the functional relevance of rs1741981. The Functional Single Nucleotide Polymorphism (F-SNP; http://compbio.cs.queensu.ca/F-SNP/) database integrates information obtained from 16 bioinformatics tools and databases about the functional effects of SNPs [27]. For rs1741981, F-SNP used TFSearch (http://www.cbrc.jp/research/db/TFSEARCH.html) [28] and Consite (http://asp.ii.uib.no:8090/cgi-bin/CONSITE/consite) [29] tools. Both tools computationally predict changes in transcription factor binding according to the allele of re1741981 (T allele vs. C allele) (Fig. 2 and Supplementary Fig. 1). F-SNP indicated 0.268 for the FS score of rs1741981. The FS score is a functional SNP scoring system provided by F-SNP to distinguish features of disease-related SNPs versus neutral SNPs [30]. The median FS score for neutral SNPs is 0.1764, whereas for disease-related SNPs, the median rises to 0.5 [30]. Thus, rs1741981 may be functionally important.
There are several potential limitations to our results. First, sufficient statistical power, conferred by a large sample size, is an important aspect of genetic studies. The small number of patients enrolled in the present study limits the scope of our results. Second, subsequent replication studies should consider as many clinical and demographic confounding factors as possible and should be performed in diverse ethnic backgrounds. Third, haplotype approaches for HDAC1 would result in more precise associations. For example, one previous study showed the advantage of examining both individual SNPs and haplotypes to further pin-point a potential causal SNP [31]. Finally, although a computational method suggested that rs1741981 may be functionally important, further mechanistic studies are needed to clarify the precise role of rs1741981 and HDAC1 in the response to corticosteroid treatment.
KEY MESSAGE
1. In the present study, we identified rs1741981 in HDAC1, which showed a significant relationship with asthma severity.
2. We also found that this genetic variation was associated significantly with lung function improvements in response to systemic corticosteroid treatment in adult asthmatics and in response to inhaled corticosteroids treatment in childhood asthmatics.
3. This is the first report of a significant association between a genetic variation in HDAC1 and the response to corticosteroid treatment in asthmatics.


 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print





